MALEZAS-RESISTENTES A HERBICIDAS: ¿ CÓMO ESTO OCURRE?
HERBICIDE-RESISTANT WEEDS: ¿HOW DOES THIS OCURR?
*Dustin Moreno-Serrano. Universidad de Panamá, Facultad de Ciencias Agropecuarias. Panamá.
dustin.moreno@up.ac.pa https://orcid.org/0000-0001-9134-2222
*Autor de Correspondencia: dustin.moreno@up.ac.pa
Recibido: 07/04/2025 Aceptado: 05/05/2025
DOI https://doi.org/10.48204/j.ia.v7n2.a7496
![]()
Resumen. Desde su introducción a mediados de la década de 1920, los herbicidas se han convertido en una herramienta fundamental para el control de malezas en los sistemas de producción agrícola modernos. Sin embargo, el uso continuo de herbicidas con el mismo sitio de acción ha promovido el desarrollo y la proliferación de malezas resistentes en los campos de cultivo. En años recientes, los esfuerzos por comprender los mecanismos de resistencia a nivel molecular se han convertido en un componente clave para prevenir la evolución de la resistencia. Tanto los mecanismos de resistencia por alteración del sitio de acción (TSR) como aquellos no relacionados con el sitio de acción (NTSR) se han identificado como las formas más comunes de resistencia a herbicidas. El TSR suele estar caracterizado por mutaciones que afectan la proteína blanco del herbicida, lo que limita su acceso al sitio de acción. La mayoría de estas mutaciones ocurren en el dominio catalítico o en sus proximidades. Además, el aumento en el número de copias del gen que codifica la proteína blanca puede estar involucrado dentro de los mecanismos de resistencia TSR. Por otro lado, NTSR abarca una gama más amplia de mecanismos de resistencia como degradación metabólica, la reducción en la absorción y translocación del herbicida. Usualmente en los mecanismos NTSR suelen estar involucradas múltiples familias de genes, que incluyen, pero no se limitan a citocromo P450, glutatión S-transferasas, entre otros. Ambos mecanismos, TSR y NTSR pueden coexistir en una misma planta, como resultado de un proceso evolutivo que conduce a la ineficacia de uno o varios herbicidas con diferentes modos de acción.
Palabras clave: Citocromo P450, glutatión S-transferasas, mecanismos de resistencia, NTSR, reducida absorción - translocación, TSR.
Abstract. Since their introduction in the mid-1920s, herbicides have become a fundamental tool for weed control in modern agricultural production systems. However, the continued use of herbicides with the same site of action has promoted the development and proliferation of resistant weeds in crop fields. In recent years, efforts to understand resistance mechanisms at the molecular level have become a key component in preventing the evolution of resistance. Both target-site resistance (TSR) and nontarget-site resistance (NTSR) mechanisms have been identified as the most common herbicide resistance mechanisms. TSR is usually characterized by mutations that affect the herbicide's target protein, limiting its access to the site of action. Most of these mutations occur either in or near the catalytic domain. Furthermore, increased copy numbers of the gene encoding the target protein and also involved in TSR resistance mechanisms. On the other hand, NTSR encompasses a broad range of resistance mechanisms, such as herbicide-enhanced metabolism, reduced uptake, and translocation. NTSR mechanisms typically involve multiple gene families, including, but not limited to, cytochrome P450, glutathione S-transferases, and others. Both TSR and NTSR mechanisms can coexist in the same plant, resulting in an evolutionary process that leads to the ineffectiveness of one or more herbicides with different modes of action.
Keywords: Cytochrome P450, glutathione S-transferases, resistance mechanisms, NTSR, reduced translocation and uptake, TSR.
INTRODUCCIÓN
En su gran mayoría, las plantas no deseadas que se encuentran en campos de cultivos agrícolas se consideran malezas, ya que interfieren y afectan un cultivo de interés. Esto ocurre no solo dificultando las labores de siembra y cosecha, sino que también compitiendo por recursos esenciales como agua, nutrientes, espacio y luz. La continua presión de malezas en la agricultura moderna conllevó la introducción de herbicidas, permitiendo a los agricultores obtener un control efectivo y económico y logrando así obtener altos rendimientos bajo diferentes escenarios. La introducción de los herbicidas revolucionó notablemente la agricultura global, al igual que el manejo de malezas por los últimos 70 años (Oerke, 2006). Sin embargo, la introducción de los herbicidas impulsó una fuerte dependencia por los agroquímicos como parte fundamental en los sistemas agrícolas, dando paso a la idea de que el control químico es la alternativa más eficaz para controlar y reducir las poblaciones de malezas.
Actualmente, el uso de herbicidas se ha convertido en la principal herramienta utilizada por los agricultores en el control de malezas. No obstante, es evidente que el incremento en el uso de herbicidas ha conllevado la constante presión de selección en plantas y, por lo tanto, la evolución y desarrollo de resistencia no solo a uno, sino a múltiples herbicidas con diferentes modos y mecanismos de acción. En la actualidad existen un gran número de clases químicas de herbicidas, las cuales actúan en más de 25 sitios de acción específicos dentro de la célula vegetal (Gaines et al., 2020). Recientemente, en el año 2024, el Comité de Acción de Resistencia a los Herbicidas (HRAC, por sus siglas en inglés) publicó la nueva clasificación de herbicidas, con el objetivo de proveer una fuente confiable de información que pueda ser empleada en el manejo de malezas, y por lo tanto retrasar la evolución de resistencia.
La resistencia a herbicidas es uno de los desafíos más significativos en la agricultura global, impactando directamente la sostenibilidad de los sistemas agrícolas. Este fenómeno ocurre cuando una población de malezas es sometida constantemente a la aplicación de herbicidas, y como resultado, evoluciona mecanismos que le permiten sobrevivir a dosis que antes eran letales. Usualmente, esto sucede donde no existe una rotación eficiente de herbicidas con diferentes sitios de acción, lo que favorece el desarrollo y la proliferación de biotipos resistentes. Estos mecanismos de resistencia se agrupan en dos grandes categorías: resistencia por alteración del sitio de acción (TSR, por sus siglas en inglés), que implica cambios en la proteína o enzima objetivo, impidiendo su acción al limitar la capacidad de unión del herbicida al sitio de acción; y la resistencia no relacionada con el sitio de acción (NTSR), que abarca un grupo más complejos de mecanismos como rápida detoxificación o metabolismo mejorado, reducción de absorción o translocación (Bo et al., 2017).
Ante esta problemática, resulta esencial comprender cómo ocurre la resistencia a herbicidas, ya que este conocimiento es fundamental para crear estrategias de manejo eficiente y sostenibles de malezas. Por lo tanto, la presente revisión de literatura busca brindar una actualización sobre los mecanismos de resistencia más comunes reportados en malezas.
MATERIALES Y MÉTODOS
La metodología de selección de la información incluyó: exploración, selección, análisis criterioso e interpretación de los datos de literatura relevante. Se priorizaron artículos experimentales que permitirán identificar los mecanismos de resistencia y sus implicaciones en el manejo de malezas en cultivos agrícolas.
Los aportes de la presente revisión brindan una comprensión integral sobre los principales mecanismos de resistencia a herbicidas en malezas reportados, así como de sus implicaciones en el manejo agronómico. Este aporte busca contribuir al desarrollo de estrategias eficientes en el manejo de malezas perniciosas, y a su vez, prolongar la evolución de resistencia, lo cual representa, sin lugar a duda uno de los desafíos para la agricultura a nivel global.
RESULTADOS Y DISCUSIÓN
Evolución de Resistencia en Malezas
Una maleza resistente es aquella planta que, mediante selección natural, ha desarrollado uno o varios mecanismos genéticos o fisiológicos que le permiten sobrevivir a la aplicación de un herbicida que, en condiciones normales, controlaría eficazmente individuos susceptibles de la misma especie. Cuando se aplica un herbicida, se crea una constante presión de selección en la que los biotipos susceptibles mueren, pero los biotipos tolerantes o resistentes sobreviven, se reproducen y transmiten sus genes de resistencia a su descendencia (Gaines et al., 2020). De esta forma, aumenta el porcentaje de resistencia en una población y al mismo tiempo hay una disminución de la susceptibilidad. Por lo tanto, si existe constante presión de selección, es muy probable que la frecuencia de individuos resistentes aumente con el tiempo, lo que dificulta el control efectivo de malezas (Damalas y Koutroubas, 2024).
Desde mediados de la década de 1920, los herbicidas se han convertido en la principal herramienta en el control de malezas en sistemas de producción agrícola (Duke, 2012). Sin embargo, el uso constante y muchas veces excesivo de herbicidas ha favorecido la aparición y propagación de malezas resistentes. Actualmente existen 534 casos únicos de resistencia reportados en diversas especies de malezas a nivel mundial, algunas de las cuales han desarrollado mecanismos que les permiten resistir a herbicidas con más de cuatro sitios de acción diferentes (Figura 1) (Heap, 2025). Por otro lado, el incremento de casos de resistencia en diferentes especies de malezas a grupos de herbicidas específicos como (inhibidores del acetolactato sintasa (grupo 2), inhibidores de la fotosíntesis en PSII (grupo 5), acetil coenzima A carboxilasa (grupo 1) e inhibidores de EPSPS (grupo 9) han aumentado con 176, 87, 51 y 60 respectivamente (Figura 2) (Heap, 2025). Esta situación es preocupante debido a la complejidad, elevado costo y largo tiempo requerido para desarrollar un nuevo herbicida con un nuevo sitio de acción diferente.
Figura 1
Aumento cronológico de casos únicos de resistencia en malezas resistentes a herbicidas en el mundo.
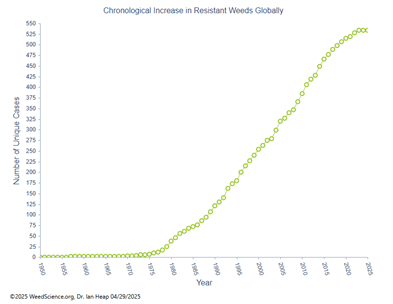
Fuente. Tomado de www.weedscience.org.
Figura 2
Aumento de especies de malezas resistentes según sitios de acción herbicida.
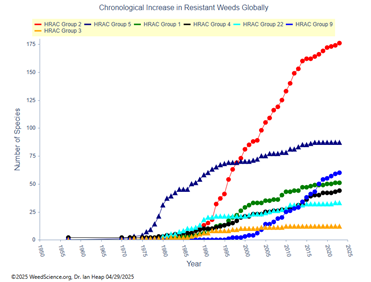
Fuente. Tomado de www.weedscience.org
La evolución de resistencia a herbicidas en malezas usualmente es gobernada por mecanismos genéticos heredables que se expresan como resultado del uso repetido de herbicidas con el mismo modo de acción. Resistencia por alteración del sitio de acción (TSR) y resistencia no relacionada con el sitio de acción (NTSR) son las dos vías en la que las malezas envuelven resistencia. Sin embargo, es importante resaltar que los mecanismos NTSR son más complejos de elucidar y también los más importantes en cuanto a manejo se refiere. Esto se debe a que generalmente están involucrados en resistencia a múltiples herbicidas con diferentes sitios de acción, lo que resulta en la ineficacia de herbicidas que incluso no habían sido usados anteriormente (Figura 3).
Figura 3
Representación esquemática de los mecanismos de resistencia a herbicidas.
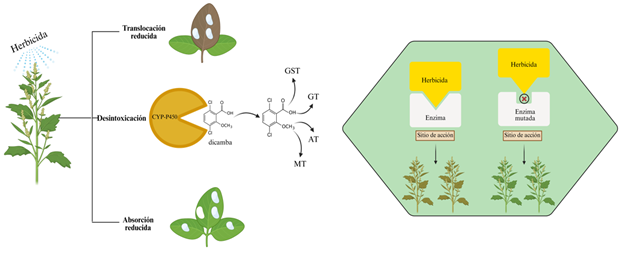
Nota. Izquierda: NTSR (resistencia no relacionada al sitio de acción). Las plantas pueden desarrollar resistencia a herbicidas al reducir su translocación, detoxificación mediante el citocromo (CYP450) (fase I) y conjugación con moléculas como glutatión-S-transferasas (GST), glucosil transferasas (GT), aminotransferasas (AT) y malonyl transferasas (MT) (fase II). Derecha: TSR (resistencia relacionada al sitio de acción). Las plantas pueden desarrollar mutaciones puntuales en el gen que codifica la enzima clave, impidiendo la unión del herbicida. Figuras creadas con BioRender.
Resistencia por alteración del sitio de acción (Target Site Resistance, TSR)
De acuerdo con nuestra revisión de literatura, en la actualidad se identifican tres tipos principales de resistencia por alteración del sitio de acción: deleción de codones, sobreexpresión del gen del sitio de acción y mutación en el sitio de accion del herbicida (Patzold et al., 2006; Gaines et al., 2010; Carvalho-Moore et al., 2022; Qi et al., 2025; Wang et al., 2025).
Deleción de codón:
En Amaranthus palmeri, se ha confirmado que la eliminación completa del codón correspondiente a la glicina en la posición 210 (ΔG210-PPO2) confiere resistencia a herbicidas que inhiben la enzima protoporfirinógeno oxidasa (PPO) (Patzoldt et al., 2006; Carvalho-Moore et al., 2022). Además, se validó que la expresión de ΔG210 en el gen PPO2 de A. palmeri, confirió una notable tolerancia a los herbicidas fomesafen y saflufenacil. Esta tolerancia fue demostrada en plantas transgénicas de arroz (Oryza sativa) y Arabidopsis thaliana, las cuales expresaron la mutación ΔG210-PPO2 (Carvalho-Moore et al., 2022). Estos resultados sugieren que la mutación ΔG210-PPO2 podría introducirse en cultivos para el desarrollo de plantas transgénicas con un grado alto de tolerancia o resistencia a herbicida del grupo de los PPO, lo cual sería de gran utilidad en el manejo de malezas, reduciendo la dependencia por los herbicidas no selectivos.
Sobreexpresión del gen del sitio de acción:
En algunos casos, la resistencia puede darse mediante una sobreexpresión del gen del sitio de acción, lo que conduce a una producción elevada de la proteína objetivo. Esto causa que una dosis letal de herbicida no sea suficiente para inhibir completamente la actividad de la enzima, permitiendo a la planta sobrevivir. El incremento en el número de copias del gen del sitio de acción, en el genoma, puede deberse a cambios en los mecanismos regulatorios que controlan su transcripción (Gaines et al., 2020). Panozzo et al., 2021 describieron el mecanismo de resistencia de dos especies de malezas comúnmente encontradas en campos de arroz en Italia, Echinochloa cruz-galli (L.) y Echninochloa oryzicola a herbicidas inhibidores del acetolactato sintasa (ALS). Los autores señalan que ambas especies contenían diferentes mutaciones en Ala122Asn y Trp574Leu para E.crus-galli y E. oryzicola respectivamente. Sin embargo, ambas mutaciones se localizaron en el gen (ALS1), para ambas especies, el cual fue significativamente sobreexpresado en comparación a las otras copias del gen ALS2 y ALS3. Estos resultados indican que los altos niveles de resistencia en ambas especies no solamente pueden deberse a mutaciones puntuales en genes específicos, sino también la sobreexpresión de dichos genes (Torra y Alcántara-De la Cruz, 2022).
Mutaciones puntuales:
Estas representan uno de los mecanismos más comunes de resistencia a herbicidas. Los primeros casos documentados de resistencia por mutaciones puntuales se registraron en malezas tratadas con herbicidas inhibidores de ALS y ACCase, donde cambios en una sola base del ADN alteraban la proteína, impidiendo la unión del herbicida. Sin embargo, en este caso, el enfoque se centrará en los herbicidas hormonales, cuyo modo de acción complejo ha dificultado la caracterización de mutaciones. En Australia, Sisymbrium orientale, se validó que una mutación en el co-receptor AUX/IAA34 (Leu175Pro) confiere altos niveles de resistencia a 2,4-D, ácido 2-metil-4-clorofenoxiacético (MCPA) y fluroxipir (Qi et al., 2025). Un caso similar fue reportado en la maleza de difícil control Kochia scoparia, donde se identificó una sustitución (GWPPV→NWPPV) en la región altamente conservada del degrón en el co-receptor AUX/IAA16, lo que conllevo al desarrollo de resistencia a dicamba, 2,4-D y fluroxipir (LeClere et al., 2018). Hasta el momento, en el grupo de los herbicidas hormonales, el degrón parece ser una parte fundamental en la estabilidad y percepción del herbicida en la planta. Por lo tanto, mutaciones en esta sección del co-receptor, conlleva al desarrollo de resistencia e insensibilidad de la planta a la hormona (Moreno-Serrano et al., 2024).
Resistencia no relacionada con el sitio de acción (Non Target Site Resistance, NTSR)
Dentro de los mecanismos NTSR, la mayoría de los artículos se enfocan en la rápida detoxificación del herbicida o en un metabolismo mejorado. Este mecanismo involucra la acción de diversas familias de enzimas y rutas metabólicas de degradación que pueden inactivar la acción fitotóxica del herbicida. A continuación, se presentarán dos contribuciones relacionadas con la familia de enzimas citocromo P450 (CYP450) y una con las glutatión S-transferasas (GST).
Citocromo P450:
En poblaciones de A. palmeri resistentes al inhibidor del hidroxifenil piruvato dioxigenasa (HPPD) tembotrion, se identificó que el incremento en el número de copias del gen CYP72A1182 permitió una detoxificación rápida de tembotrion a hidroxi-tembotrion, el cual mostró no ser fitotóxico (Rigon et al., 2025). Además, se hipotetizó que el incremento de la expresión génica de CYP72A1182 puede estar regulado tanto por elementos del Cis en la región promotora del gen como regulación de los factores de transcripción. Por otra parte, un caso similar fue reportado en Lolium rigidum donde el incremento de copias del gen CYP81A10v7 confirió alta y moderada resistencia a múltiples herbicidas de diversos modos de acción. Estos incluyen herbicidas no solo del grupo de los inhibidores del acetil-coenzima A carboxilasa (ACCase)-(diclofop-metil y tralcoxidim), acetolactato sintasa (ALS)-(clorsulfurón) y hidroxifenil piruvato dioxigenasa (HPPD)-(tembotrion) sino también a los inhibidores del fotosistema II (PSII)-(atrazina y clorotolurón) e inhibidores de la tubulina (MA)-(trifluralina) (Han et al., 2021).
Glutatión-S-transferasas:
Después de las enzimas CYP, el segundo grupo más importante corresponde a las GST, las cuales están involucradas en la fase II de detoxificación de xenobióticos. Este grupo de enzimas tienen la capacidad de conjugar glutatión, con el herbicida o con productos intermedios de la fase I mediada por CYP, transformándolos en compuestos no fitotóxicos, lo que facilita su transporte y compartimentalización dentro de la célula vegetal (Gaines et al., 2020). Actualmente, se ha identificado que las glutatión S-transferasas (GST) más abundantes y asociadas a la resistencia a herbicidas pertenecen a las clases tau (U) y phi (F), las cuales están presentes tanto en monocotiledóneas como en dicotiledóneas. En Echinochloa glabrescens, se reportó recientemente, por primera vez, resistencia a inhibidores de la acetil-coenzima A carboxilasa (ACCase), específicamente al herbicida metamifop, mediante un mecanismo de rápida detoxificación mediado por la acción de glutatión S-transferasas (GST) (Li et al., 2023). Mediante análisis por LC-MS/MS, los autores identificaron que el metamifop podía ser rápidamente degradado a metabolitos no fitotóxicos, como N-(2-fluorofenil)-2-(4-hidroxifenoxi)-N-metilpropionamida y 4-(6-clorobenzo[d]oxazol-2-iloxi)fenol.
Otros mecanismos relacionados NTSR:
Algunos mecanismos de resistencia no relacionados con el sitio de acción (NTSR) y que son poco comunes incluyen procesos como la reducida translocación y absorción. En Plantago lanceolata fue validado que, la reducida translocación era el principal mecanismo de resistencia contra el herbicida inibidor del fotosistema I (PSI)-(paraquat) (Ndou et al., 2024). Sin embargo, en Conyza bonariensis se demostró que el mecanismo de resistencia al herbicida inhibidor del (EPSPS)-glifosato era reducida translocación al sitio de acción (Palma-Bautista et al., 2021). Los autores determinaron, mediante el uso de glifosato marcado con ¹⁴C, que la absorción en plantas sensibles fue superior en más de un 25 % en comparación con las plantas resistentes.
CONCLUSIONES
Los mecanismos de resistencia a herbicidas tanto aquellos que involucran directamente el sitio de acción (TSR) como los que no involucran directamente el sitio de acción (NTSR), son fundamentales para comprender como las plantas desarrollan diversos mecanismos que les permiten sobrevivir a agentes xenobióticos como los herbicidas.
En el contexto actual, donde el uso de herbicidas sigue siendo fundamental, es crucial comprender los complejos mecanismos de resistencia para desarrollar herramientas de diagnóstico rápido. Esto permitiría la implementación temprana de estrategias en el manejo integrado de malezas y ayudaría a retrasar la evolución de la resistencia, garantizando la sostenibilidad de los sistemas agrícolas a largo plazo.
Bo, A. B., Won, O. J., Sin, H. T., Lee, J. J. y Park, K. W. (2017). Mechanisms of herbicide resistance in weeds. Korean Journal of Agricultural Science, 44(1), 1-15.
Carvalho-Moore, P., Rangani, G., Langaro, A. C., Srivastava, V., Porri, A., Bowe, S. J., Lerchl, J. y Roma-Burgos, N. (2022). Field-evolved ΔG210-ppo2 from Palmer Amaranth confers pre-emergence tolerance to PPO-inhibitors in Rice and Arabidopsis. Genes, 13(6), 1044.
Damalas, C. A. y Koutroubas, S. D. (2024). Herbicide resistance evolution, fitness cost, and the fear of the superweeds. Plant Science, 339, 111934.
Duke, S. O. (2012). Why have no new herbicide modes of action appeared in recent years?. Pest management science, 68(4), 505-512.
Gaines, T. A., Duke, S. O., Morran, S., Rigon, C. A., Tranel, P. J., Küpper, A. y Dayan, F. E. (2020). Mechanisms of evolved herbicide resistance. Journal of Biological Chemistry, 295(30), 10307-10330.
Gaines, T. A., Zhang, W., Wang, D., Bukun, B., Chisholm, S. T., Shaner, D. L., Nissen, S., Patzoldt, W., Tranel, P., Culpepper, S., Grey, T., Webster, T., Vencill, W., Sammons, D., Jian, J., Preston, C. y Westra, P. (2010). Gene amplification confers glyphosate resistance in Amaranthus palmeri. Proceedings of the National Academy of Sciences, 107(3), 1029-1034.
Rigon, C. A., Küpper, A., Sparks, C., Montgomery, J., Peter, F., Schepp, S., Perez-Jones, A., Tranel, P., Beffa, R., Dayan, F. E. y Gaines, T. A. (2025). Function of Cytochrome P450 CYP72A1182 in Metabolic Herbicide Resistance Evolution in Amaranthus palmeri Populations. Journal of Experimental Botany, eraf114.
Han, H., Yu, Q., Beffa, R., González, S., Maiwald, F., Wang, J. y Powles, S. B. (2021). Cytochrome P450 CYP81A10v7 in Lolium rigidum confers metabolic resistance to herbicides across at least five modes of action. The Plant Journal, 105(1), 79-92.
Heap, I. (2024). International herbicide-resistance weed database. https://www.weedscience.org/Home.aspx Accesado en mayo 2025.
LeClere, S., Wu, C., Westra, P. y Sammons, R. D. (2018). Cross-resistance to dicamba, 2, 4-D, and fluroxypyr in Kochia scoparia is endowed by a mutation in an AUX/IAA gene. Proceedings of the National Academy of Sciences, 115(13), E2911-E2920.
Li, Q., Zhao, N., Jiang, M., Wang, M., Zhang, J., Cao, H. y Liao, M. (2023). Metamifop resistance in Echinochloa glabrescens via glutathione S‐transferases‐involved enhanced metabolism. Pest Management Science, 79(8), 2725-2736.
Moreno-Serrano, D., Gaines, T. A. y Dayan, F. E. (2024). Current Status of Auxin‐Mimic Herbicides. Outlooks on Pest Management, 35(3), 105-112.
Ndou, V., Kotze, D., Marjanovic-Painter, B., Phiri, E. E., Pieterse, P. J. y Sonopo, M. S. (2024). Reduced Translocation Confers Paraquat Resistance in Plantago lanceolata. Agronomy, 14(5), 977.
Oerke, E. C. (2006). Crop losses to pests. The Journal of Agricultural Science, 144(1), 31-43.
Palma-Bautista, C., Vázquez-García, J. G., Domínguez-Valenzuela, J. A., Ferreira Mendes, K., Alcántara De la Cruz, R., Torra, J. y De Prado, R. (2021). Non-target-site resistance mechanisms endow multiple herbicide resistance to five mechanisms of action in Conyza bonariensis. Journal of Agricultural and Food Chemistry, 69(49), 14792-14801.
Panozzo, S., Mascanzoni, E., Scarabel, L., Milani, A., Dalazen, G., Merotto, A., Tranel, P. y Sattin, M. (2021). Target-site mutations and expression of ALS gene copies vary according to Echinochloa species. Genes, 12(11), 1841.
Patzoldt, W. L., Hager, A. G., McCormick, J. S. y Tranel, P. J. (2006). A codon deletion confers resistance to herbicides inhibiting protoporphyrinogen oxidase. Proceedings of the National Academy of Sciences, 103(33), 12329-12334.
Qi, Y., Krishnan, M., Tucker, M. y Preston, C. (2025). A point mutation in IAA34 confers resistance to the auxin herbicide 2, 4‐D in Sisymbrium orientale. Pest Management Science.
Torra, J. y Alcántara-De la Cruz, R. (2022). Molecular mechanisms of herbicide resistance in weeds. Genes, 13(11), 2025.
Wang, J., Du, Y., Zhang, L., Deng, Y., Wang, T., Wang, S. y Ji, M. (2025). Pro‐197‐Ser mutation combinations in acetolactate synthase (ALS) homoeologous genes affect ALS inhibitor herbicide resistance levels in Monochoria korsakowii. Pest Management Science, 81(4), 1894-1902.